
Mines Chimie PC 2024
| Thème de l'épreuve | Synthèse totale de l'aigialomycine D. Le dioxygène et la respiration branchiale chez le requin. |
| Principaux outils utilisés | cinétique chimique, thermodynamique, chimie organique, diffusion |
| Mots clefs | solubilisation, myoglobine, hémoglobine, respiration |
Corrigé
:👈 gratuite pour tous les corrigés si tu crées un compte
👈 l'accès aux indications de tous les corrigés ne coûte que 1 € ⬅ clique ici
👈 gratuite pour tous les corrigés si tu crées un compte
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
👈 gratuite pour ce corrigé si tu crées un compte
- - - - - - - - - - - -
Énoncé complet
(télécharger le PDF)















Rapport du jury
(télécharger le PDF)




Énoncé obtenu par reconnaissance optique des caractères
A2024 --- CHIMIE PC
Cmb
Concours commun
Mines-Ponts
ÉCOLE DES PONTS PARISTECH,
ISAE-SUPAERO, ENSTA PARIS,
TÉLÉCOM PARIS, MINES PARIS,
MINES SAINT-ÉTIENNE, MINES NANCY,
IMT ATLANTIQUE, ENSAE PARIS,
CHIMIE PARISTECH - PSL.
Concours Mines-Télécom,
Concours Centrale-Supélec (Cycle International).
CONCOURS 2024
Durée de l'épreuve : 4 heures
L'usage de la calculatrice et de tout dispositif électronique est interdit.
ÉPREUVE DE CHIMIE
Durée de l'épreuve : 4 heures
L'usage de la calculatrice et de tout dispositif électronique est interdit.
Les candidats sont priés de mentionner de façon apparente
sur la première page de la copie :
CHIMIE - PC
L'énoncé de cette épreuve comporte 16 pages de texte.
Si, au cours de l'épreuve, un candidat repère ce qui lui semble être une erreur
d'énontcé, il le signale sur sa copie et
poursuit sa composition en expliquant les raisons des initiatives qu'il est
amené à prendre.
Les sujets sont la propriété du GIP CCMP. Ils sont publiés sous les termes de
la licence
Creative Commons Attribution - Pas d'Utilisation Commerciale - Pas de
Modification 3.0 France.
Tout autre usage est soumis à une autorisation préalable du Concours commun
Mines Ponts.
Ce problème comporte deux parties indépendantes intitulées « Synthèse totale de
l'aigialomycine D »
et « Le dioxygène et la respiration branchiale chez le requin ».
Les données utiles à la résolution du problème sont fournies à la fin de
l'énoncé (pages 14, 15 et 16).
Synthèse totale de l'aigialomycine D
L'aigialomycine D (Figure 1), un macrolide résorcinolique à 14 chainons, a été
isolé à partir du
champignon de mangrove Aigialus parvus. Ce composé fait partie d'une grande
famille de produits
naturels qui possèdent une structure de macrolide à 14 chaînons fusionnée à une
unité benzénoïde. Les
macrolides résorcinoliques présentent un fort potentiel thérapeutique. Par
exemple, 1l a été montré que
l'aigialomycine D possédait une très forte activité antipaludique (IC : 6,6
ug/mL contre Plasmodium
falciparum) ainsi qu'une cytotoxicité importante contre le carcinome
épidermoïde humain (1C5o : 3,0
ug/mL contre les cellules KB).
OH O
(@)
HO [OS
OH
OH
aigialomycine D
Figure I. Structure de l'aigialomycine D.
Ces propriétés biologiques intéressantes ont stimulé les travaux de synthèse
orientés vers les macrolides
résorcinoliques et leurs analogues. Une synthèse de l'aigialomycine D (Figure
1), publiée par le groupe
de Danishefsky en 2004, utilise comme produit de départ le
2-désoxy-D-ribopyranose (1).! Ce composé
existe en solution aqueuse sous trois formes différentes (Schéma 1) : la forme
ouverte minoritaire
(environ 1 %), et deux formes cycliques comportant respectivement un cycle à 6
atomes (environ 75 %)
et un cycle à 5 atomes (environ 24 %).
O0. .OH Of
CT -- Ho CRE 2-désoxy-D-ribofuranose
HO ; OH (24 %)
OH
2-désoxy-D-ribopyranose (1) 2-désoxy-D-ribose
(75 %) (1 %)
Les pourcentages des différentes formes sont donnés à 25 °C.
Schéma I. Différentes formes du substrat de départ I en solution aqueuse.
Q1- Représenter tous les stéréoisomères du 2-désoxy-D-ribose (forme ouverte) et
donner les
relations de stéréoisomérie entre chaque paire de structures.
Q2- Représenter la forme cyclique à 5 atomes appelée 2-désoxy-D-ribofuranose,
en indiquant sans
ambigüité la position des différents groupements par rapport au plan du cycle.
Combien de
stéréoisomères du 2-désoxy-D-ribofuranose existe-t-1l en solution ?
l X. Geng, S. J. Danishefsky, Org. Lett. 2004, 6, 413-416.
Page 1/16
Q3- Quelle réaction permet de passer de la forme linéaire à la forme
2-désoxy-D-ribopyranose ?
Proposer un mécanisme en milieu acide pour cette transformation.
L'alcyne 8, un précurseur de l'aigialomycine D, a été préparé à partir du
2-désoxy-D-ribopyranose (1).
Sa synthèse débute par deux réactions qui conduisent successivement aux
composés 3 et 4 (Schéma 2).
HO
O .OH O OH --
CT Réactifs 2 CT -- PPhs à PivCI
HO o L O Et;N, DMAP
OH 77 4 K
1 3
1) SNS |
ZT 2NBr NN PivO
1) Hydroboration SO3-pyr THE, 0 °C
2) Oxydation DMSO, EN 2) Hydrolyse HO O
f
O°C.1h 8 OK
O à
PivCI = > 0 DMAP = ven pyr = 4 }
DMSO = 5-0 THE = d ]
Schéma 2. Synthèse du précurseur 8 de l'aigialomycine D.
Q4- Proposer des réactifs (notés réactifs 2) et des conditions réactionnelles
pour préparer
efficacement le composé 3 à partir du composé 1.
Le composé 4, mis en présence de chlorure de pivaloyle (PivCI ou chlorure de
2,2-diméthylpropanoyle)
avec de la triéthylamine et de la N N-diméthylpyridin-4-amine (DMAP) dans le
dichlorométhane,
conduit à la formation du composé 5. Ce dernier subit une séquence
hydroboration-oxydation pour
donner le composé 6 (Schéma 2).
Q5- Donner la structure du composé 5 et proposer un mécanisme justifiant sa
formation à partir du
composé 4 (sans faire intervenir la DMAP dans le mécanisme).
Q6- Donner la structure du composé 6 et préciser l'ensemble des réactifs
nécessaires à l'obtention
de ce dernier à partir du composé 5. Justifier la régiosélectivité de la
réaction.
Le composé 6 est alors oxydé en aldéhyde 7 via une réaction de Parikh-Doering"
(Schéma 2).
| Q7- Donner la structure de l'aldéhyde 7.
Le traitement du composé 7 par un organozincique, un organométallique dont la
réactivité est analogue
à celle d'un organomagnésien mixte, tout en étant moins nucléophile et moins
basique, conduit à la
formation de l'alcyne vrai 8 après hydrolyse (Schéma 2).
Q8- Donner deux raisons pour lesquelles 1l n'est pas possible d'utiliser
l'analogue
organomagnésien (HC=CCH2MzgBr) de l'organozincique impliqué dans cette
transformation (le pK:
2 J.R. Parikh, W. E. Doering, J. Am. Chem. Soc. 1967, 89, 5505.
Page 2/16
d'un alcyne vrai est de l'ordre de 25). Par analogie avec la réactivité des
organomagnésiens mixtes,
proposer un mécanisme rendant compte de la transformation de l'aldéhyde 7 en
alcyne 8.
L'alcyne 8 est ensuite transformé en l'acide carboxylique 10 en plusieurs
étapes, avec l'obtention
intermédiaire du composé 9 (Schéma 3).
HO,C
NS NS K
NS PivO SN K NS K
HO D + TBSO Do + TBSO O
8 KR 9 KR
O
Piv = x TBS = BuMe,sSi
Schéma 3. Formation de l'acide carboxylique 10 à partir de l'alcyne 8.
Q9- Avec l'aide du Document |, proposer un enchainement de plusieurs réactions
afin d'obtenir
le composé 9 à partir de l'alcyne 8. On justifiera l'ordre des réactions et les
éventuels problèmes
rencontrés. Les réactifs principaux ainsi que la structure des composés
intermédiairement obtenus
seront précisés sans donner les mécanismes des différentes transformations.
Document I. Protection de la fonction alcool.
Dans une synthèse, les groupements hydroxyles sont, la plupart du temps,
protégés. Parmi les
groupements protecteurs couramment utilisés, on rencontre les éthers silylés
(R-OSiR'R°R*) qui sont
formés via des réactions de substitution.
R1
N 1
Re sic Ro
/ J --
RS Si 3
R-O > _R-O RO ---- R-OH
Et;N
éther silylé
Les éthers silylés sont retirés en présence d'ions fluorure (comme Bu4NF), afin
de régénérer la
fonction alcool. Parmi les éthers silylés courants on rencontre le
fert-butyldiméthylsilyle (R' = Bu,
R° = R° = Me) abrégé TBS, le triéthylsilyle (R' = R° = R° = Et) abrégé TES ou
le triméthylsilyle (R'
= R°= R° = Me) abrégé TMS.
Le groupement MOM (méthoxyméthyle) est également un groupement protecteur très
utile en
synthèse. La fonction alcool est protégée en présence de MOMCI en milieu
basique (Et3N), et ce
groupement protecteur peut être retiré en milieu acide aqueux.
MOMCI R H
ON EN 9 OMe Ho
(R-OMOM)
| Q10- Proposer des conditions (réactifs et solvants) pour former l'acide 10 à
partir du composé 9.
Quelques étapes, non décrites 1c1, ont ensuite permis d'obtenir le diène 11 à
partir de l'acide 10. Par
réaction de métathèse cyclisante en utilisant le catalyseur de Grubbs de
seconde génération, le diène 11
a conduit au macrolide à 14 atomes 12 (Schéma 4).
Page 3/16
cataylseur
de Grubbs 11 ©
O NS
x CH,Cl
TBSO O OTBS
11 12
Schéma 4. Obtention du macrolide 12 par métathèse cyclisante.
Q11- Seul le stéréoisomère de configuration Æ£ du composé 12 est obtenu lors de
cette réaction.
Quelle méthode expérimentale a permis de confirmer la configuration Æ de la
double liaison formée
? Justifier votre réponse.
Dans la publication décrivant cette synthèse,' les auteurs indiquent que le
composé 12 est un mélange
de deux stéréoisomères, notés 12a et 12b, pour lesquels les valeurs suivantes
sont rapportées dans la
partie expérimentale :
12a : [a] =--124,6 (c 0,17, CHCH) et 12b : [a] = 173,3 (c 0,41, CHCH)
Q12- À quelle grandeur correspondent ces résultats ? Commenter les valeurs
obtenues. La présence
de deux stéréoisomères pose-t-elle un problème pour la suite de la synthèse ?
La réaction de fermeture aboutissant au squelette macrocyclique à 14 atomes de
l'aigialomycine D a
constitué un vrai défi synthétique, et de nombreux groupes de recherches se
sont attelés à cette tâche.
Outre la méthode de métathèse cyclisante utilisée par le groupe de
Danishefsky,' une méthode
stéréosélective de cyclisation (au niveau de la double liaison carbone-carbone
créée), utilisant une
catalyse par un complexe du nickel, a été développée par le groupe de
Montgomery (Schéma 5).°
MOMO O MOMO O
Lo L.Ni(O) cat.
(L= PPh3)
D
>
Et:SiH MOMO
13 OTBS
Schéma 5. Cyclisation catalysée par un complexe du nickel. °
Le mécanisme de cette cyclisation comporte quatre étapes (une partie des
structures a été simplifiée
pour ne laisser apparents que les groupes fonctionnels réagissant).
3 C. C. Chrovian, B. Knapp-Reed, J. Montgomery, Org. Lett. 2008, 10, 811-814.
Page 4/16
" 2) L_Ni(O) s)
14 --
Schéma 6. Mécanisme de la cyclisation catalysée par un complexe du nickel.
Q13- Donner le nom des deux étapes A et D de ce mécanisme.
Q14- Donner les variations du nombre d'oxydation du métal nickel dans chacune
des quatre étapes.
La suite de la synthèse de l'aigialomycine D consiste en une réaction de
Diels-Alder entre la triple liaison
carbone-carbone du composé 12, précédemment décrit (Schéma 4), avec le diène
15. Le produit
intermédiairement formé 16 (non isolé) subit, dans les mêmes conditions, une
réaction de rétro-Diels-
Alder pour conduire au composé 17 où les groupements triméthylsilyles (TMS) ont
été ôtés lors du
traitement de la réaction (Schéma 7 et Document 1 page 3).
OTMS
140 °C
---- [16] --
15 TBSO O
17 OR
Schéma 7. Formation du composé 17 via des réactions de Diels-Alder et
rétro-Diels-Alder.
Q15- Donner la structure du composé intermédiaire 16 formé et justifier la
régiosélectivité de la
réaction à l'aide des données du Tableau 1.
Page 5/16
Tableau 1. Caractéristiques des orbitales frontalières de molécules modèles des
réactifs 12 et IS.
Coefficient sur l'atome n°
. 6 Orbitale
OK nn. 1 2 3 + 5 6 7
°| | HO (-10,7 eV) 0,50 --0,05 -0,63 0,49 0,30 0,10 --0,09
4
u BV (0,1 eV) 0,38 --0,55 0,22 0,66 -0,19 0,16 ---0,06
. Coefficient sur l'atome n°
OMe Orbitale
; A, 1 2 3 + 5 6 7 8
eo 8 HO (-8,1 eV) 0,23 0,43 0,39 0,33 0,59 0,17 -0,18 0,24
e
5 BV (0,6 eV) 0,16 -0,62 0,33 0,37 -0,55 0,09 0,16 0,14
Q16- Donner la structure du composé organique, possédant quatre atomes de
carbone, formé lors de
la réaction de rétro-Diels-Alder.
Le diène impliqué dans la réaction de Diels-Alder, le
5,5-diméthyl-1,3-bis(triméthylsilyloxy)cyclohexa-
1,3-diène 15, a été préparé en quelques étapes à partir du composé commercial
18 (Schéma 8).
O EtO,C" "CO.Et
20
HOT KOH HCI aq.
18 > > [21] ---- 22 ---- 23 24
EtONa, EtOH 80 °C 100 °C
19 2 h, 80 °C C43H2505 C:44H4604
OTMS
TMSCI
ZnCb, Et3N TMSO
15
Schéma 8. Synthèse du 5,5-diméthyl-1,3-bis(triméthylsilyloxy)cyclohexa-I,
3-diène IS.
L'oxyde de mésityle 19 (4-méthylpent-3-én-2-one) est obtenu par condensation
d'un précurseur
organique à trois atomes de carbone 18 en milieu basique. Il est ensuite traité
par le malonate de diéthyle
20 en présence d'éthanolate de sodium dans l'éthanol pour conduire à un produit
intermédiaire 21 de
formule brute C13H205 qui cyclise rapidement en 22, de formule brute C:1H1604.
Le composé 22 est
ensuite chauffé au reflux en présence d'une solution aqueuse d'hydroxyde de
potassium pour conduire
au composé 23 qui subit une décarboxylation (perte d'une molécule de dioxyde de
carbone) en milieu
acide à chaud pour former le composé 24. Une dernière étape permet de former le
diène 15 attendu.
Q17- Donner la structure du composé organique 18 (à trois atomes de carbone)
précurseur de
l'oxyde de mésityle 19. À quel type de réaction correspond cette transformation
?
Q18- Donner la structure de l'intermédiaire 21 formé par condensation, sous
contrôle orbitalaire, du
diester malonique 20 sur l'oxyde de mésityle 19. Expliquer la régiosélectivité
de la formation de ce
composé à l'aide des données du Tableau 2.
Page 6/16
Tableau 2. Caractéristiques des orbitales frontalières du composé 19.
Coefficient sur l'atome n°
Orbitale
5 Of! | 2 3 4 5 6 7
EUR 7 HO (-9,9 eV) 0,52 --0,10 --0,66 ---0,42 0,24 0,24 0,06
BV (0,1 eV) --0,37 0,56 0,16 --0,66 0,18 0,18 -0,15
Q19- Donner la structure du composé cyclique 22 ainsi que le mécanisme de sa
formation, sachant
que l'intermédiaire 21 (non isolé) et le produit 22 sont obtenus dans les même
conditions.
Les données spectroscopiques de RMN 'H du composé 22 sont les suivantes :
RMN 'H (400 MHz, CDCHk) : ô 4,22 (q, J = 8,0 Hz, 2H), 3,63 (s, 2H), 2,89 (s,
1H), 2,32 (s, 2H), 1,21
(t, J = 8,0 Hz, 3H), 0,99 (s, 6H).
(s = singulet ; d = doublet ; {= triplet ; g -- quadruplet)
Q20- Attribuer les signaux relevés sur le spectre de RMN 'H du composé 22.
Q21- Quelles bandes caractéristiques peut-on attendre en spectroscopie
infrarouge pour ce composé
22 ?
Q22- Donner les structures des composés 23 et 24.
L'aigialomycine D est finalement obtenue en quelques étapes (Schéma 9) à partir
de l'intermédiaire 17
(Schéma 7). Les deux groupements hydroxyles résorcinoliques du composé 17 sont
d'abord protégés
par le groupement méthoxyméthyle (MOM) pour former le composé 25, avant de
créer la double liaison
carbone-carbone de type styrène du composé 26. Une dernière étape permet alors
d'obtenir
l'aigialomycine D.
--+ aigialomycine D
Schéma 9. Fin de la synthèse de l'aigialomicine D.
Q23- Proposer un mécanisme pour la formation de 25 à partir de 17 (Document 1
page 3). Donner
le nom des fonctions créées lors de cette étape.
Q24- Proposer un schéma réactionnel, en plusieurs étapes, expliquant la
formation le composé 26 à
partir de 25. Justifier la régiosélectivité de la réaction de formation de la
double liaison styrénique.
Q25- Proposer des conditions réactionnelles pour la formation de
l'aigialomycine D à partir de 26.
Page 7/16
Le dioxygène et la respiration branchiale chez le requin
Le requin respire grâce à la présence de cinq à sept fentes branchiales sur les
côtés de sa tête. Lorsqu'il
se déplace, l'eau de mer pénètre par sa bouche, passe sur les branchies
constituées d'un grand nombre
de lamelles, puis ressort par ses fentes branchiales. Le dioxygène dissous dans
l'eau de mer se fixe au
moment du contact avec les capillaires sanguins des branchies. Ce mode de
respiration contraint le
requin de haute mer à nager en permanence pour que l'eau traverse en continu
ses branchies. S1 le requin
s'arrête de nager, c'est l'asphyxie. C'est pourquoi lorsqu'un requin est pris
dans un filet de pêche, 1l
meurt asphyxié. La vitesse moyenne d'un requin est de l'ordre de 8 km/h, mais
les plus rapides d'entre
eux, comme le requin Mako, peuvent atteindre, sur de courtes distances, des
vitesses allant jusqu'à 70
km-h".
Ce problème comporte quatre parties. Les trois premières abordent des points
importants pour la
modélisation de la respiration branchiale du requin traitée dans la dernière
partie.
Solubilité du dioxygène dans l'eau de mer
Le requin puise le dioxygène nécessaire à sa survie dans son milieu
environnant. Il est donc important
de connaitre la concentration en dioxygène dissous dans l'eau de mer. Des
observations ont montré
qu'elle dépend d'un grand nombre de paramètres, comme les variations de
température, les courants
marins, la consommation du dioxygène par des bactéries aérobies, la
photosynthèse, la salinité, la pluie,
le vent... mais aussi de la pression atmosphérique. Dans la suite du problème,
par souci de
simplification, l'eau de mer sera assimilée à de l'eau pure.
Q26- À l'aide des données fournies (annexes page 14), déterminer la
concentration en dioxygène
dissous à la surface de la mer dont l'eau est à 25 °C et est soumise à une
pression de Ï bar.
Q27- Comment varie la solubilité du dioxygène quand la température du milieu
augmente ? Justifier
en détaillant le déplacement d'équilibre. Estimer la variation relative (en
pourcentage) de la solubilité
du dioxygène s1 la température augmente de 6 °C.
En laboratoire, des études cinétiques montrent que le transfert moléculaire du
dioxygène de l'air vers de
l'eau pure (choisie comme modèle pour l'analyse) suit une loi du premier ordre
de la forme suivante :
-- = +k,(c-- co)S (1)
où dan est la quantité de matière de dioxygène qui se dissout dans l'eau
pendant le temps dt, S représente
la surface d'échange, c est la concentration en dioxygène dissous dans l'eau à
un instant t, c, correspond
à la solubilité du dioxygène à la température considérée et k, est le
coefficient de transfert de masse.
| Q28- Préciser le signe de l'expression de dn/dt et l'unité du coefficient k,.
Des mesures expérimentales du coefficient k, ont donné les relevés suivants en
fonction de la
température T (Tableau 3).
Page 8/16
Tableau 3. Coefficient k, en fonction de la température T.
T (°C) 17 25 35 40 50
k,(SL) 4,0-104 4,4104 49-104 5,1:104 5,6:104
S1 l'on trace la courbe Ink; = f(1/T) on obtient une droite de pente --950 et
d'ordonnée à l'origine --
4,5.
Q29- En déduire une estimation de l'énergie d'activation E, de la réaction de
dissolution du
dioxygène gazeux dans l'eau.
La saturation en dioxygène x est définie comme le rapport de la concentration
en dioxygène sur sa
solubilité.
Q30- Estimer la durée nécessaire à une eau à 17 °C à 90 % de saturation sur 10
centimètres de
profondeur pour retrouver une saturation x de 99 % (la concentration en
dioxygène sera supposée
égale en tout point de la couche d'eau considérée). Commenter cette valeur.
Transport et stockage du dioxygène chez le requin
Chez le requin l'apport de dioxygène au niveau des muscles est assuré par deux
protéines, l'hémoglobine
et la myoglobine. Ces protéines peuvent fixer réversiblement le dioxygène en le
complexant sur l'atome
de fer d'un groupement (appelé hème) qu'elles portent. La myoglobine ne possède
qu'un seul hème,
alors que l'hémoglobine dispose de quatre hèmes, chacun étant susceptible de
fixer une molécule de
dioxygène. L'hémoglobine est impliquée dans le transport du dioxygène, tandis
que la myoglobine
intervient dans les processus de stockage. On considérera dans cette partie que
l'équilibre de dissolution
du dioxygène décrit en début de problème, s'applique entre l'air et le milieu
biologique. On considère
la réaction de complexation de la myoglobine (Mb) en oxymyoglobine (MbO:) dont
la constante de
formation est f :
MD(ag) * O2(ag = MbO; (0)
Le pourcentage de saturation Y,, de la myoglobine est défini comme le rapport
entre la concentration
d'oxymyoglobine et la concentration totale en myoglobine (libre et complexée
par Où) :
[MbO; |
Ymb =
[MbO; | + [Mb]
P
Q31- Montrer que le pourcentage de saturation YL, peut s'écrire sous la forme :
Yyp = =. =
O5 TE Mb
Exprimer la constante P\y, en fonction des données. Que représente Pyp ?
Pour l'hémoglobine, la réaction de complexation avec le dioxygène, de constante
de formation f,, est
la suivante :
Hb(ag) + N O2(ag = Hb(O2} (20)
Q32- Montrer que le pourcentage de saturation de l'hémoglobine Y,, peut
s'écrire sous la forme :
PO)
= ñ =:
P6 + Pb)"
Yub
Page 9/16
I Exprimer la constante Py, en fonction des données.
YHb
--YHb
Des mesures expérimentales montrent que le tracé de in en fonction de InPo, est
une droite de
Y Mb
1--Y Mb
également d'obtenir les valeurs des constantes Pur et Ps, respectivement égales
à 30 et 7 mbar.
pente 3 et que le tracé de in en fonction de inP,, est une droite de pente 1.
Ces mesures permettent
Q33- Les valeurs des coefficients directeurs des régressions linéaires
étaient-elles attendues ?
Expliquer.
La pression en dioxygène diminue le long de la circulation sanguine des artères
jJusqu'aux organes. Au
niveau des capillaires sanguins la pression en dioxygène est d'environ 130 mbar
alors qu'elle se réduit
à 40 mbar au niveau des muscles.
Q34- Justifier l'intérêt des deux modes de transport du dioxygène et préciser
l'équation chimique
de transfert du dioxygène de l'hémoglobine vers la myoglobine.
Production de l'énergie chez le requin
En biologie, la respiration cellulaire peut être représentée par la réaction
d'oxydation du glucose
(C6H1206 ay) qui sert de combustible. Le dioxygène transporté par l'hémoglobine
ou stocké par la
myoglobine sert de comburant à la réaction.
Q35- Déterminer l'enthalpie standard de la réaction de combustion complète AH?
du glucose à 298
K.
Q36- Calculer la constante thermodynamique de la réaction de combustion
complète du glucose à
298 K. Commenter.
La respiration cellulaire n'oxyde pas le glucose en une seule étape. Sa
dégradation suit une série de
réactions, chacune catalysée par une enzyme. Des atomes d'hydrogène sont
extraits du glucose lors de
certaines étapes, mais 1ls ne sont pas transférés immédiatement au dioxygène.
Ils vont passer par un
intermédiaire organique nommé nicotinamide adénine dinucléotide ou NAD" qui
joue le rôle de receveur
d'électrons et de protons en formant NADH. L'ensemble du processus biochimique
est inscrit dans un
cycle catalytique, il est donc nécessaire de régénérer les accepteurs NAD° qui
ont été réduits en
composés NADH.
Q37- Combien d'électrons sont nécessaires à l'oxydation totale d'une molécule
de glucose ? Estimer
le potentiel standard du couple glucose/CO. Combien de molécules de NAD° sont
alors impliquées
dans le cycle biologique simplifié à compléter et reproduire sur la copie
(Figure 2) ?
CeH1206 | | | |
Figure 2. Cycle biologique simplifié d'oxydation du glucose.
Le bilan énergétique de l'oxydation complète du glucose fait apparaître la
synthèse de 36 molécules
d'ATP (indispensable à l'effort musculaire) par phosphorylation d'ADP. L'ATP
est utilisé dans tous les
Page 10/16
processus cellulaires nécessitant de l'énergie. Cette molécule sert de stockage
de l'énergie puisque son
hydrolyse libère environ 30 kJ:mol ".
Q38- En déduire la valeur du rendement énergétique du mécanisme de respiration
cellulaire par
oxydation du glucose. Commenter.
En fait l'alimentation d'un requin est pauvre en carbohydrates comme le
glucose, mais riche en acide
gras. Les lipides constituent ainsi la principale source d'énergie métabolique
chez les requins et leur
consommation est étroitement liée aux conditions environnementales et aux
cycles biologiques, tels que
ceux liés à l'alimentation, à la reproduction et à la migration. La lipolyse
est la réaction de dégradation
des lipides mise en oeuvre pour fournir de l'énergie chez le requin. Les acides
gras possèdent de longues
chaînes carbonées dont la formule brute est approchée à (CH).
Q39- Donner l'équation de la réaction de combustion complète des acides gras en
fonction de n,
puis calculer la valeur de l'enthalpie de réaction A_HY correspondante en
fonction de n.
Q40- Comparer les énergies libérées par les réactions de combustion du glucose
et des acides gras,
en se ramenant à une mole de dioxygène pour chaque réaction. Commenter.
La vitesse de déplacement du requin
Dans le modèle étudié ci-dessous, le requin est assimilé à un moteur ditherme
fonctionnant entre une
source froide (la mer) et une pseudo source chaude dont l'énergie libérée par
mole de dioxygène EUR, est
estimée à 0,4 MJ-mol !. Pour les applications numériques, on considère un
requin de 100 kg et un
rendement énergétique de 40 %.
Q41- Les besoins en dioxygène d'un requin sont caractérisés par le flux molaire
en dioxygène ® (en
mole-s '). Exprimer ® en fonction de la masse du requin m, de sa puissance
massique P,,, de &. et
du rendement 7 de la machine ditherme que constitue le requin, puis en donner
une application
numérique (le rendement d'un moteur ditherme est le rapport du travail fourni à
l'extérieur et de
l'énergie thermique fournie au système par la source chaude).
Quand un requin respire, le transfert du dioxygène dissous dans l'eau de mer
vers le sang s'effectue au
niveau des membranes des lamelles branchiales. En régime stationnaire, on
considère un modèle de
conduction particulaire axiale selon l'axe Ox à travers les membranes des
lamelles de surface S (Figure
3). On note [O>]|h la concentration en dioxygène dans l'eau de mer et [O2],
celle dans le sang du requin.
O2 (9)
[O2]
[O2] = #S
=
mer = sang x
Figure 3. Modèle simplifié de conduction
particulaire au niveau des branchies du requin.
Page 11/16
Pour étudier ce modèle, on utilise la loi de Fick (Figure 4) qui traduit un
lien de proportionnalité entre
la cause de la diffusion selon l'axe Ox (inhomogénéité de la concentration
particulaire) et la conséquence
(flux molaire ® à travers une surface S). Dans cette équation, c représente la
concentration en particules
et D le coefficient de diffusion, qui est constant. Le signe « -- » indique que
le flux de particules a lieu
dans le sens des concentrations décroissantes (évolution spontanée donc
irréversible, dans le sens de la
tendance à l'homogénéisation).
Figure 4. Loi de Fick.
Q42- Exprimer le flux molaire @ en dioxygène à travers les lamelles en fonction
de [O{s, [O2]m, S
la surface des lamelles, e l'épaisseur de la membrane des lamelles, et D le
coefficient de diffusion
du dioxygène dans les lamelles.
La fixation du dioxygène par l'hémoglobine du sang étant très efficace, 1l est
raisonnable de supposer
que la concentration en dioxygène libre dissous dans le sang est négligeable.
| Q43- En déduire une expression de d en fonction de la pression partielle en
dioxygène de l'air.
On considère en première approximation que le transport du dioxygène nécessaire
à la respiration est
limité par sa diffusion à travers les lamelles.
Q44- Donner une estimation de la surface des lamelles dans ce modèle qui ne
tient compte que de
la diffusion interne dans les lamelles.
En réalité la surface des lamelles branchiales d'un requin de 100 kg est
d'environ $ = 30 m°. La
différence observée avec le résultat du calcul précédent s'explique par le fait
que le modèle simplifié
(Figure 3) néglige le transport externe convectif du dioxygène de l'eau de mer
vers les lamelles
branchiales. Aussi, un modèle plus réaliste de la conduction introduit la
concentration en dioxygène à
la surface des lamelles [O2], qui est distincte de la concentration en
dioxygène dans l'eau de mer [O2]»
(Figure 5).
[Obls
[O2]m = [O2],
mer
lamelle
sang
X
Figure 5. Modèle de conduction particulaire
tenant compte du transfert convectif du dioxygène
marin.
Le flux molaire de transfert convectif du dioxygène marin vers les lamelles, à
travers une surface S
s'écrit de façon analogue à l'équation (1), page 8 :
P = +ke(1O2lm -- [O2]z)S
Dans cette équation, le coefficient de transfert de masse k; dépend du
coefficient de diffusion du
dioxygène dans l'eau de mer D,,, de la viscosité cinématique de l'eau v, de la
vitesse du fluide + et
d'une longueur caractéristique L (L est 1c1 la longueur d'une lamelle, L = 2
mm) :
Page 12/16
k, -- DER g 2
Q45- En traduisant la continuité du flux molaire de dioxygène, déterminer la
vitesse minimale de
nage d'un requin de 100 kg lui garantissant un flux suffisant en dioxygène pour
sa respiration.
Commenter.
Fin de l'épreuve
Page 13/16
Annexes
Annexe I. Constantes usuelles et approximation de calculs.
Constante de Faraday : F % 10° C-mol!.
Constante des gaz parfaits R = 8 J-K-1:mol-1.
Il sera considéré que RTIn10 = 6 kJ-mol-1 à 298 K ; e! = 3 ; In10 & 2,3.
Annexe 2. Données thermodynamiques (considérées indépendantes de la
température).
Espèce CO> (g) H20 (1) CéH1206 (CH)n O> (g)
(aq)
AFH° (kJ-mol ") | -390 -280 --1300 --30n 0
S9 (J-mol !'-K_ | 210 70 210 210
ll
)
Annexe 3. Potentiels standards d'oxydoréduction à DH = 0 et à 298 K (par
rapport à E.S.H.).
E(H°/EH; (9) = 0,0 V.
E9(O; (HO) -- 1,23 V.
Annexe 4. Constantes physicochimiques.
Masses molaires (g-mol '): C=12;0=16;H=1.
Viscosité cinématique de l'eau à 20 °C : v = 10 °m°s |.
Coefficients de diffusion du dioxygène :
- dans les tissus du requin : D = 2,0:10-9 m°.s !
- dans l'eau de mer : D,, = 10-° m°°s !
La constante thermodynamique (K7) associée à l'équilibre de dissolution du
dioxygène gazeux :
O2 (g) ---- O2 (aq)
Ky = 1,3-10-3 et A,.H° = 12 kJ-mol-1 à 298 K.
Annexe 5. Autres données.
+ On considère que l'air est composé, en fractions molaires, de 80 % de diazote
et 20 % de dioxygène.
+ Les besoins énergétiques (puissance massique) d'un requin sont estimés à P,,
= 0, 6 W-kg !.
° Les branchies du requin sont constituées d'un grand nombre de lamelles entre
lesquelles l'eau de mer
circule. En progressant le long des lamelles l'eau s'appauvrit en oxygène au
profit du sang qui circule à
contre-courant à l'intérieur des lamelles. Au sein de chaque lamelle,
l'épaisseur de la membrane entre
l'eau et le sang est d'environ e = 15 um.
+ La vitesse moyenne de nage des requins est de 8 km/h. Lorsqu'ils chassent,
les plus rapides atteignent
des vitesses allant jusqu'à 70 km/h.
Page 14/16
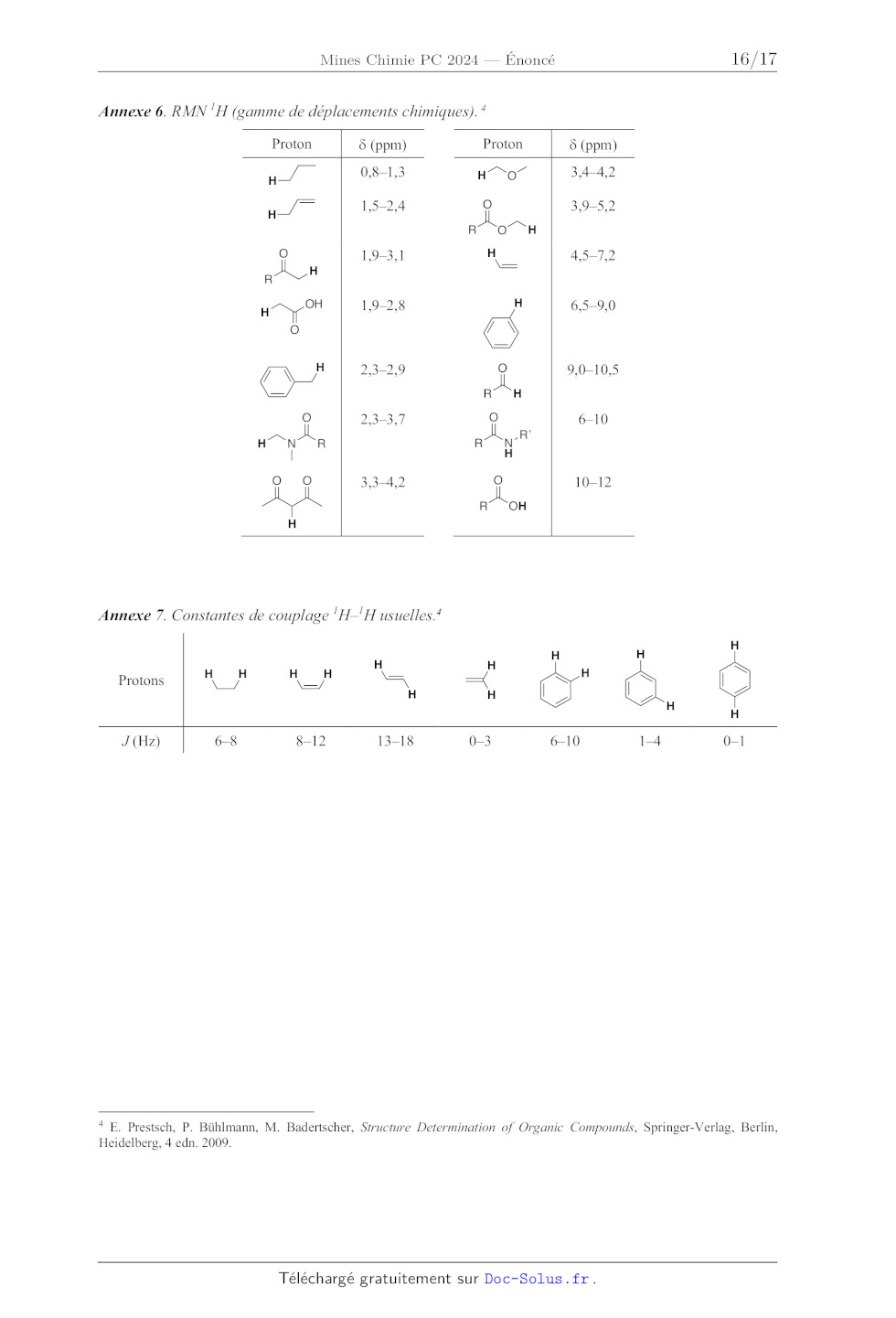
Annexe 6. RMN 'H (gamme de déplacements chimiques). *
Proton Ô (ppm) Proton Ô (ppm)
_ NAT
u/-- 0,8-1,3 Ho 3,4-4,2
-- : O _
4/-- 1,5-2,4 ? 3,9-5,2
R7 OT
O 1,9---3,1 H 4,5--7,2
XCH D D \ -- 9 7,
R
"0 ul 1,9-2,8 H 6,5-9,0
j CO
a 2,3--2,9 ï 9,0-10,5
R H
O 2,3-3,7 e) 6--10
ete. a ye
| H
O oO 3,342 O 10-12
AS à oH
H
Annexe 7. Constantes de couplage H--'H usuelles."
H H H H A H H
Protons \ _/ \__/ = -- b! C
H H
H
J (Hz) 68 8-12 13-18 0-3 6-10 1-4 0-1
* E. Prestsch, P. Bühlmann, M. Badertscher, Sfructure Determination of Organic
Compounds, Springer-Verlag, Berlin,
Heidelberg, 4 edn. 2009.
Page 15/16
Annexe 8. Table infrarouge (IR) des nombres d'onde de quelques groupes
fonctionnels.*
Liaison Nombre d'onde (cm ')
O-H libre 3580-3670
O-H alcool lié 3200-3400
N-H amine, imine 3100-3500
N-H amide 3100-3500
C-H alcyne (=C-H) 3300-3310
C-H alcène (--C-H) 3000-3100
C-H aromatique 3030-3080
C-H alcane (-C-H) 2800-3000
C-H aldéhyde 2750-2900
O-H acide carboxylique lié 2500-3200
C=C 2100-2250
C=N 2120-2260
C=O anhydride 1700-1840
C=O chlorure d'acyle 1770-1820
C=O ester 1735-1750
C=O acide carboxylique 1700-1725
C=O amide 1640-1670
1650-1730
C=O aldéhyde et cétone
C=C
C=C aromatique
N=0O
C=N
C-O
C-F
C-CI
C-Br
C-I
abaissement de 20 à 30 cm !
Si Conjugaison
1625-1685
1450-1600
1510-1580
1600-1680
1050-1450
1000-1040
700-800
600-750
500-600
Page 16/16